|
|
Die Bedeutung der biologischen
Verwandtschaft und Herkunft der bestatteten Individuen in megalithischen
Gesellschaften Antoinette Goujon |
|
2 Prähistorische Anthropologie 2.2.1. Die genetischen Untersuchungen in Irland 2.3.3 Die Isotopen-Untersuchungen in der Schweiz 3.1 Das kulturelle Gedächtnis
nach Assmann |
Zusammen mit
dem assoziierten archäologischen Kontext, ethnoarchäologischen Analogien und
Ansätzen der sozialen Anthropologie und Kulturwissenschaften ergibt sich
anhand der Ergebnisse bioarchäologischer Untersuchungen eine Möglichkeit, die
komplexen und vielfältigen Gesellschaften Europas zur Zeit des Neolithikums
sowohl auf biologischer wie auch auf sozialer Ebene zu untersuchen. Die hier
vorgestellten Beispiele der in megalithischen Anlagen bestatteten Individuen
neolithischer Gesellschaften zeigen auf, dass die biologische Verwandtschaft
(biological kinship)
einen nicht zu unterschätzenden Anteil an der sozialen Organisation hatte;
allerdings spielte auch die nicht-biologische Verwandtschaft (social kinship) eine erhebliche Rolle. Biologische und soziale
Reproduktion sind in den Megalithen-erbauenden Gesellschaften des
europäischen Neolithikums unweigerlich miteinander verbunden. Diese
Verknüpfung diverser Erkenntnisse zeigt auf, dass in der Erforschung der
Megalithen-erbauenden Gesellschaften des europäischen Neolithikums noch viele
Fragen offen und viele interdisziplinäre Ansätze möglich sind. Seit der Mitte
des 19. Jahrhunderts ist der Begriff der Megalithik
(gr. mégas ‹gross›, gr. lithos ‹Stein›), genauer dessen Adjektiv ‹megalithisch›,
im britischen und französischen Raum gebräuchlich. Schliesslich
wurde am internationalen Kongress der Anthropologie und Prähistorischen
Archäologie (Congrès internationale d’anthropologie et d’archéologie
préhistorique, CIAAP) im Jahr 1867 in Paris
beschlossen, ‹keltische Monumente› von nun an ‹megalithische Monumente› zu
nennen, und sie von Monumenten wie Obelisken und Zyklopenmauern abzugrenzen
(vgl. Boulestin 2016: 62f). Megalithische Monumente
können einzelne Menhire, Steinkreise und -reihen, Dolmen, Gang-, Pass- sowie
auch Steinkistengräber sein, die im europäischen Raum während des
Neolithikums von ca. 4500 bis 2500 v. Chr. erbaut und genutzt wurden (vgl.
Schwegler 2016: 33–149). Allerdings zählen auch historische und
zeitgenössische ethnologische Beispiele aus megalithischen Gesellschaften zur
Kategorie der megalithischen Monumente, beispielsweise die Bauten der Toraja und Niha auf den Inseln
Indonesiens (vgl. Steimer-Herbet 2018: 69–76,
86–97), die Grabstätten der Merina auf Madagaskar
oder die ahu-Plattformen und berühmten moai-Statuen der Osterinsel, Überreste der
verschwundenen Rapanui (vgl. Cauwe 2014:
321–330; Cauwe 2016: 221–225). Die Frage, die
sich Bruno Boulestin stellt, was denn nun den ‹Megalithismus› ausmacht und beinhaltet, ist also nicht so
leicht, wenn überhaupt, zu beantworten; der Begriff, der gegen Ende des 19.
Jahrhundert auftaucht, ist bis heute in Gebrauch, und erhielt im Laufe der
Zeit die verschiedensten Definitionen (vgl. Boulestin
2016: 62f). Dabei handelt es sich beim ‹Phänomen des Megalithismus›
um eine grobe Vereinfachung der komplexen Strukturen und Gesellschaften, die
mit megalithischen Monumenten in Verbindung gebracht werden. Nach Jean
Leclerc (1931–2012), der sich seit den 1960er-Jahren mit den Bestattungsriten
und Kollektivbestattungen des Neolithikums im französischen Raum befasste und
die französische archéologie funéraire
massgeblich prägte, müsse man innerhalb des
gebräuchlichen Begriffs des Megalithismus’ drei Phänomene
unterscheiden: den wahren Megalithismus (mégalithisme proprement dit), den Monumentalismus (monumentalisme) und die Bestattung in Kollektivgräbern (l’inhumation en sépulture collective, vgl. Boulestin
2016: 57). Allerdings ist auch die Verknüpfung der Monumentalität (oder deren
Abwesenheit) in Bezug auf die Kollektivbestattungen des Neolithikums mit den
bekannten ethnologischen Vergleichen essentiell, um einen Einblick in die
Gesellschaften zu erhalten, die mit megalithischen und teilweise auch nicht-megalithischen
Bauten assoziiert werden, wenn man sich die Frage stellt, um was für
Gesellschaften es sich im Neolithikum handelte, die diese megalithischen
Bauten errichteten. Seit einigen
Jahrzehnten kommen nebst den archäologischen Strukturen und den ethnoarchäologischen
Analogien die bioarchäologischen Analysen hinzu, die wichtige Informationen
zu den bestatteten Individuen und Populationen liefern und sowohl den
materiellen und als auch den sozial-anthropologischen Kontext erweitern. Der
Begriff der Population sollte je nach Kontext und Fragestellung der
Untersuchung genauer definiert werden; die biologische Population
unterscheidet sich beispielsweise von der humanbiologischen Population. So
können mit Population auch eine kleinere Gruppe verbundener Individuen
(soziale Zusammengehörigkeit) oder auch mehrere Gruppen als eine Gemeinschaft
gemeint sein. Die technischen
und wissenschaftlichen Fortschritte in den Bereichen der prähistorischen
Anthropologie ermöglichen es, konkrete Aussagen und neue Ansätze zu Fragen
der Herkunft, Verwandtschaft und Ernährungsweise (prä-)historischer
Individuen und Populationen zu formulieren (vgl. Alt u. Vach 1994: 56f; Grupe et al. 2012: 74f, 77f; Grupe
et al. 2015: 1–3). Die Verknüpfung der bioarchäologischen Erkenntnisse mit
dem archäologischen und ethnoarchäologischen Kontext sowie Ansätze der
sozialen Anthropologie ermöglichen es, die Lebensgeschichte und -umstände
vergangener Gemeinschaften auf der individuellen Ebene als auch auf der Ebene
der Population zu rekonstruieren (vgl. Le Roy 2018: 189ff; Meyer et al.
2012a: 11f). 2 Prähistorische Anthropologie Die Methoden
und Konzepte der prähistorischen Anthropologie befassen sich mit den
menschlichen Überresten vergangener Epochen und Populationen. Bereits aus
einer ersten osteologischen Untersuchung des Skeletts können das Alter, das
biologische Geschlecht sowie etwaige Pathologien und Traumata erkannt und die
Körperhöhe geschätzt werden. Innerhalb einer Kollektiv- oder
Mehrfachbestattung kann anhand der vorhandenen Knochen die Mindestindividuenzahl (MIZ) geschätzt werden, was
zusammen mit biologischem Geschlecht und Alter eine Einschätzung der Gruppengrösse sowie der Geschlechts- und Altersverteilung
innerhalb der untersuchten Population zulässt (Paläodemographie). Ein
Beispiel dafür ist die Arbeit von Perréard Lopreno (2014: 33–36), die versuchte, die soziale Gruppengrösse der mindestens 126 im Dolmen M XII der
Fundstelle «Petit-Chasseur» (Sion, Wallis) bestatteten Individuen, die über
einen Zeitraum von ca. 500 Jahren beigesetzt wurden, zu rekonstruieren.
Allerdings ist bei einer demographischen Rekonstruktion zu beachten, dass die
Geschlechts- und Altersbestimmung erhaltungs- oder altersbedingt nicht immer
möglich ist, da beispielsweise die fragilen Knochen von Neugeborenen und
Kleinkindern schnell degenerieren oder aber nicht vorhanden sind, da
bestimmte Altersklassen oder Individuen nicht am selben Ort und auf eine
andere Art und Weise bestattet wurden. Für genetische
Analysen mittels aDNA (ancient
desoxyribonucleic acid)
und Isotopenuntersuchungen zur Herkunft, Mobilität und Ernährung, werden
Proben aus Knochen und Zähnen entnommen. Dabei ist zu beachten, dass das
passende Gewebe für die jeweilige Analyse beprobt wird, da nur so
aussagekräftige Erkenntnisse aus den Ergebnissen gezogen werden können.
Knochen und Dentin bestehen zu 70% aus einer mineralischen Komponente
(Bioapatit) sowie zu 21% aus Kollagen und 1% nicht-kollagenen Proteinen.
Zahnschmelz dagegen besteht zu mehr als 96% aus Bioapatit und enthält kein Kollagen
(vgl. Grupe et al. 2015: 63, Tab. 4.2). Um
Kontaminationen und weitere Umwelteinflüsse zu untersuchen sowie Vergleichs-
und Referenzdaten zu erhalten, werden auch aus der unmittelbaren und
entfernteren Umgebung der Überreste Proben entnommen, die zur Rekonstruktion
archäologischer Isoscapes (isotopic
landscapes) und Klimabedingungen relevant sind.
Ausschlaggebend ist besonders auch der Erhaltungszustand des untersuchten
Gewebes, weshalb eine genaue Betrachtung der taphonomischen
Umstände sowie etwaiger diagenetischer Veränderungen miteinbezogen werden
sollten (vgl. Grupe et al. 2012: 96–98, 138f; Meyer
et al. 2012b: 143). Bei der
Auswertung der bioarchäologischen Daten sollte jedoch unbedingt darauf
geachtet werden, welche Aussagen anhand der Datengrundlage überhaupt
getroffen werden können, da «[…] technisch exakte
Messwerte, die an einem bioarchäologischen Fund erhoben wurden, auf
interpretativer Ebene häufig nur Wahrscheinlichkeitscharakter haben […] Es
handelt sich um Annäherungen an die ehemalige Realität.» (Grupe
et al. 2015: 432). Meist bleiben den Archäolog*innen nur mehr Knochen und Zähne von
bestatteten Individuen erhalten, in seltenen Fällen ausgezeichneter Erhaltungsbedingungen,
wie im Permafrost oder in sehr trockenen Gebieten, allerdings auch weitere
Gewebe wie Haare, Haut, Muskelfasern oder Koprolithen. Doch auch nur anhand
von skelettalen Überresten lassen sich viele Informationen über die
Lebensumstände der einzelnen Individuen und Populationen generieren. Nebst
molekulargenetischen und biochemischen Analysen (s. unten) können erste
osteologische Betrachtungen bereits Aussagen über das biologische Geschlecht,
Alter und Körpergrösse liefern. Besonders für eine
demographische Rekonstruktion der Gruppengrösse
sind diese Informationen hilfreich. Alter und Geschlecht sind jedoch
grundlegend für weitere Analysen und Vergleiche. Das biologische Geschlecht lässt
sich mit geschultem Auge und Erfahrung bei erwachsenen Individuen mit bis zu
95% Wahrscheinlichkeit anhand des Beckens, des Schädels und der Langknochen
bestimmen (vgl. Grupe et al. 2015: 254–265; Ferembach et al. 1979: 7–45). Je vollständiger das
Skelett, desto sicherer ist die Bestimmung. Da die skelettalen Ausprägungen
von genetischen Komponenten als auch von umweltbedingten Einflüssen abhängen,
besteht eine grosse Variabilität besonders auch im
populationsspezifischen Geschlechtsdimorphismus, weshalb eine klare
Einteilung in weibliche und männliche Individuen sehr vorsichtig und
bestenfalls mit einiger Erfahrung angegangen werden sollte (vgl. Grupe et al. 2012: 104). Nicht klar bestimmbare
Individuen werden besser als solche gekennzeichnet, als dass man sie nach
Gefühl einteilt, was die Interpretation erschweren kann. Besonders bei
Kindern und Jugendlichen, deren Skelette noch nicht vollständig ausgewachsen
und oft sehr schlecht erhalten sind, ist die Geschlechtsbestimmung schwierig
oder teils gar unmöglich. Das
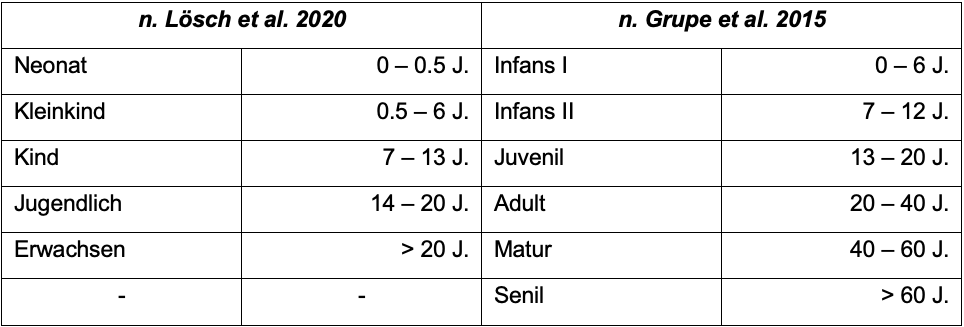
biologische Alter wird meist in Alterskategorien angegeben, die dem
jeweiligen Fundkomplex angepasst werden können (s. Tab. 1),
da genaue Zahlen schwer zu schätzen sind und das biologische Alter selten mit
dem chronologischen Alter des Individuums übereinstimmt. Das ungefähre Alter
eines Individuums kann anhand der verschiedenen Apophysen-, Epiphysenfugen-
und Schädelnahtverschlüsse (vgl. Grupe et al. 2015:
271, Abb. 8.12; 277, Abb. 8.16), dem Relief der Schambeinsymphyse sowie der
Robustheit der Knochen bestimmt werden, sowie bei Individuen bis ca. 12
Jahren anhand der durchbrochenen oder eben noch nicht durchbrochenen Zähne
(vgl. Grupe et al. 2015: 265–279; AlQahtani et al. 2010: 482, Tab. 1; Acsádi
u. Neméskeri 1970: 110–137). Bei vollständig
erwachsenen Individuen ist aber, anders als das biologische Geschlecht, das
Alter aufgrund von Belastungsveränderungen, Krankheiten, abrasiver Ernährung
und Zahnproblemen weniger genau bestimmbar.
Tab. 1: Altersklassen in Kategorien und ungefähren
Jahresangaben (nach Lösch et al. 2020: 205; Grupe
et al. 2015: 267, Tab. 8.2) Besonders
bei Kollektiv- und Mehrfachbestattungen stellt sich die Frage nach der
Verwandtschaft der Bestatteten; dies meist nicht nur innerhalb einer
einzelnen Fundstelle, sondern auch zwischen verschiedenen Fundstellen eines
Gebiets (vgl. Grupe et al. 2012: 154). Die Analyse
der noch im Knochenkollagen vorhandenen DNA ermöglicht es, den Antworten auf
diese Forschungsfragen näher zu kommen. Je nach Fragestellung und
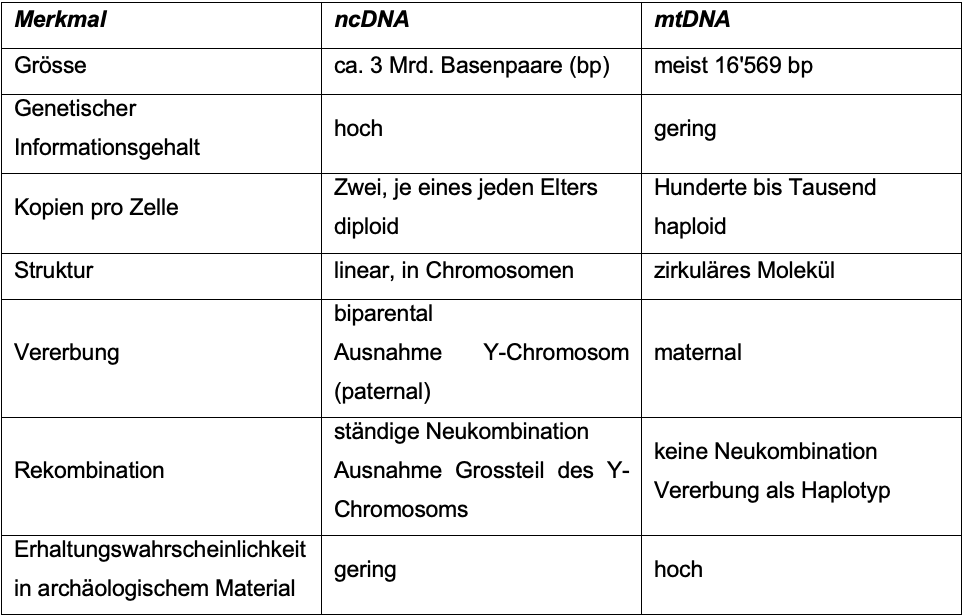
Erhaltungszustand wird die in grösserer Menge
vorhandenen mitochondriale DNA (mtDNA) oder aber die nukleäre DNA untersucht
(s. Tab. 2).
Anhand der mtDNA können matrilineare Verbindungen sichtbar werden,
während die ncDNA anhand des Y-Chromosoms Hinweise
auf patrilineare Verbindungen geben kann, aber auch enger verwandte
Individuen aufzeigen kann, wenn viele Übereinstimmungen zwischen den Genomen
bestehen (vgl. Grupe et al. 2012: 157f; Grupe et al. 2015: 515f). Ausserdem
kann die osteologische Geschlechtsbestimmung mithilfe genetischer
Untersuchungen auch bestätigt oder aber in Frage gestellt werden.
Tab. 2: Eigenschaften der ncDNA
und mtDNA im Vergleich (n. Grupe et al. 2015: 484,
Tab. 11.2) Da es sich bei aDNA um DNA handelt, die meist mehrere Jahrtausende alt
ist, gestaltet sich die Analyse jedoch etwas komplexer als bei moderner DNA.
Im Gegensatz zu moderner DNA ist aDNA stark
fragmentiert und biochemisch verändert und degradiert. Die sehr kurzen
Fragmente (<50bp) sind selten im Vergleich zur omnipräsenten
Kontamination, die aus bakterieller, fungaler,
moderner und ‹Umwelt› DNA bestehen kann (vgl. Grupe
et al. 2012: 154f; Grupe et al. 2015: 489–493).
Somit ist eine aDNA-Analyse erhaltungsbedingt nicht
mit allen Überresten durchführbar; besonders bei Altgrabungen und
Archivmaterial ist die Problematik der Kontamination sehr gross
(vgl. Grupe et al. 2015: 497, Tab. 11.5). Dem
untersuchten Gewebe, meist Knochen oder Dentin, werden wenige µg Proben
entnommen, die speziell aufbereitet und dekontaminiert werden. Vervielfältigt
wird die aDNA durch die Polymerasekettenreaktion (polymerase chain reaction, PCR). Schliesslich
gibt es mehrere Verfahren, die gewünschte aDNA-Fragmente
oder aber das gesamte Genom zu sequenzieren. Seit einigen
Jahrzehnten werden vermehrt genetische Analysen auch an prähistorischen
Überresten durchgeführt. Alt et al. (2016) publizierten eine ausführliche
Studie zur (nicht-)megalithischen Kollektivbestattung von Alto de Reinoso in Burgos, Spanien. Die Grabanlage, die
vermutlich aus einer hölzernen Totenhütte mit Steinpackung bestand, stammt
aus dem späten Neolithikum (3770–3539 v. Chr.) und wurde vermutlich über 3
Generationen genutzt (ca. 60–80 Jahre). Es lassen sich 2 Nutzungsphasen
unterscheiden; eine dritte aus der Bronzezeit wurde durch moderne
Landwirtschaft praktisch zerstört. Die aDNA-Analyse
ergab, dass sich in der unteren Schicht einige Individuen befinden, die
mütterlicherseits eng miteinander verwandt waren. Die paternale
Verwandtschaft lässt sich leider nicht genauer analysieren, da die Individuen
nicht auf Y-chromosomale Haplogruppen untersucht
wurden. Dies liegt vermutlich an den schlechteren Erhaltungschancen der ncDNA im Gegensatz zur mtDNA. Die mit der Verbreitung des ‹Megalithischen
Phänomens› immer wieder assoziierte ‹Atlantische Fassade› wurde von
Sánchez-Quinto et al. (2019) untersucht. Die Studie zeigt auf, dass in vielen
Megalithanlagen biologische Verwandtschaftsverbindungen ersichtlich sind; im
Falle Irlands sogar über mehrere Anlagen verteilt (s. unten). Sie suggeriert
darüber hinaus eine genetisch nachgewiesen patrilineare Organisation der
Gruppen, wobei sie auch darauf hinweisen, dass deutlich mehr Männer als
Frauen in den untersuchten Anlagen bestattet und analysiert wurden. Auch ist
eine Vermischung zwischen mesolithischen Jäger-und-Sammler-Populationen und
neolithischen Ackerbauern genetisch sichtbar. Die Ergebnisse sollten jedoch
kritisch betrachtet werden, da die Voraussetzungen und Probengrösse
für konkrete, allgemeine Aussagen nicht ausreichen. Sie zeigen allerdings
auf, dass die genetisch nachweisbare Komponente der Verwandtschaft in
megalithischen Gruppen durchaus eine Rolle spielte; welche genau kann daraus
jedoch nicht abgeleitet werden, da es sich hier bloss
um Ansätze handelt, die in Zukunft weiter untersucht werden müssen 2.2.1 Die genetischen Untersuchungen in Irland In
Irland wurden zur Zeit des Neolithikums (ca. 3700–2500 v. Chr.) mehrere grosse und imposante megalithischen Monumente erbaut, die
auch für Kollektivbestattungen genutzt wurden. Dabei handelt es sich um
Ganggrabkomplexe (passage tomb
complexes), Hofgräber (court
tombs) und Portalgräber (portal
tombs), aber auch um Steinkistengräber (in älterer
Literatur als Dolmen bezeichnet) des Typs Linkardstown
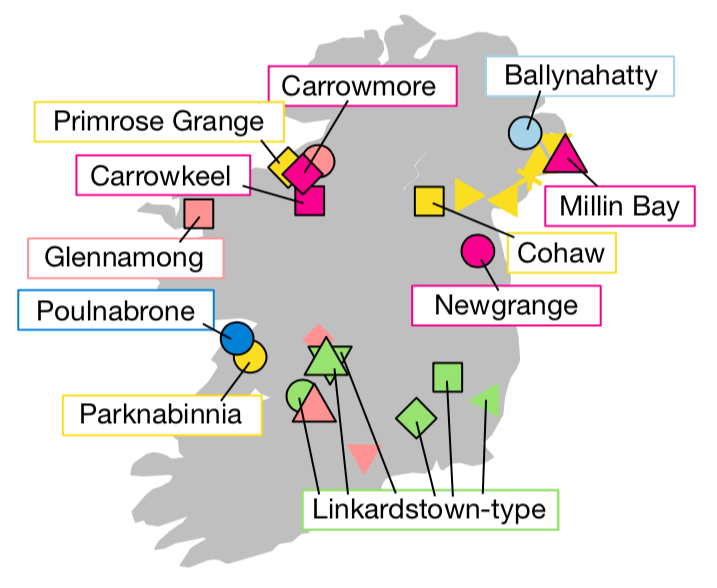
und natürliche Bestattungen ohne megalithische Bauten (s. Abb. 1).
Besonders die riesigen Ganggrabkomplexe von Newgrange
(s. Abb. 2),
Carrowkeel, Carrowmore
und Millin Bay werden als ostentative Monumente für
den öffentlichen Konsum gesehen, die auch für eine bestimmte Gruppe von
Individuen für Feste und Riten zugänglich waren, die vermutlich auch darin
bestattet wurden; Cassidy et al. argumentieren mit einer «dynastischen Elite»
(Cassidy et al. 2020: 384). Solche grossen,
intentionell erbauten Monumente, die für Kollektivbestattungen genutzt
werden, sind in vielen Regionen der Welt charakteristisch für
«multi-generational, kinship-based, corporate, aristocratic groups» (Weiss-Krejci 2018:
110), wobei der Begriff kinship, meist als ‹Verwandtschaftsbeziehungen›
übersetzt, grundsätzlich sowohl biologische als auch soziale Verbindungen
beschreibt (vgl. Weiss-Krejci 2018: 119).
Abb.
1: Fundstellen megalithischer und nicht-megalithischer Bestattungen in
Irland. Pink = Passage tomb complexes, gelb = Court tomb, blau = Portal tomb, grün
= Linkardstown-Typ, rosa = natürliche Bestattung,
hellblau = nicht klassifizierter Megalith (Cassidy et al. 2020, 385, Abb. 1c)
Abb. 2:
Newgrange Ganggrab-Komplex
(Cassidy et al. 2020, 386, Abb. 2a) Im Ganggrab von Newgrange in Irland konnten Cassidy et al. (2020) durch genetische
Analysen von insgesamt 44 Genomen (37 davon männlicher Individuen)
feststellen, dass es sich bei einem, an wichtiger Stelle im Ganggrab,
bestatteten Individuum (NG10) um einen adulten Mann handelt, der aus einer
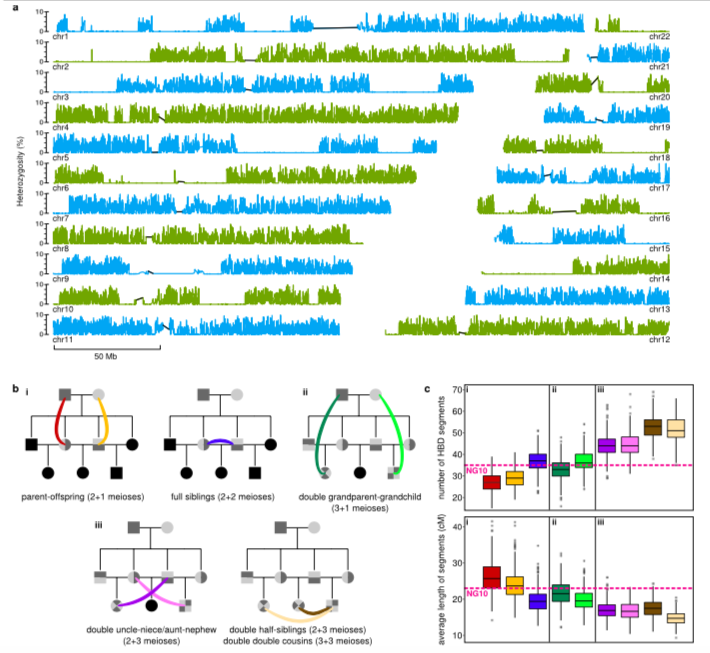
inzestuösen Verbindung 1. Grades abstammt (s. Abb. 3).
Solche inzestuösen Verbindungen stellen in den meisten Gesellschaften ein grosses soziales Tabu dar, und ist erst ein weiteres Mal
im Kontext einer megalithischen Bestattung in Schweden bekannt (vgl. Cassidy
et al. 2020: 385f). Allerdings sind einige Beispiele solcher Verbindungen
bekannt, die von der jeweiligen Gesellschaft abgesegnet wurden, meist im
Rahmen polygyner, dynastischer Eliten. Beispiele dafür wären die Gott-Könige
(deification of the ruling dynasty),
die aus den Königreichen der Inka und dem antiken Ägypten bekannt sind, sowie
in den Gesellschaften Hawaiis vor der Kolonialisierung. Auch in complex chiefdoms und early states sind solche
(Halb-)Geschwister-Heiraten bekannt, die zur Legitimierung der Macht und der
Hierarchie dienen und meist mit monumentaler Architektur einhergehen; ein
Phänomen, dass auch bei den Inka und im alten Ägypten sichtbar ist (vgl.
Cassidy et al. 2020: 385).
Abb. 3: a) Heterozygositäts-Plot für NG10, b) Verbindungsszenarien
für die hohe Homozygosität in NG10, c) Anzahl und
durchschnittliche Länge der homozygoten Segmente im Vergleich zu NG10
(Cassidy et al. 2020, Extended Data, Abb. 3) Verwandte
dieses Individuums (NG10) wurden in mehreren Kilometern Entfernung in
weiteren Grabkomplexen wie Carrowkeel (CAK532,
CAK533), Carrowmore (car004) und Millin Bay (MB6) gefunden. Anhand der Y-Chromosom-Haplogruppen wurde ersichtlich, dass es sich vermutlich
um eine identity by descent gehandelt haben könnte, die in den mittel- bis
spätneolithischen Gesellschaften Irlands eine grosse
Rolle spielte und patrilinear weitervererbt wurde. Die in den
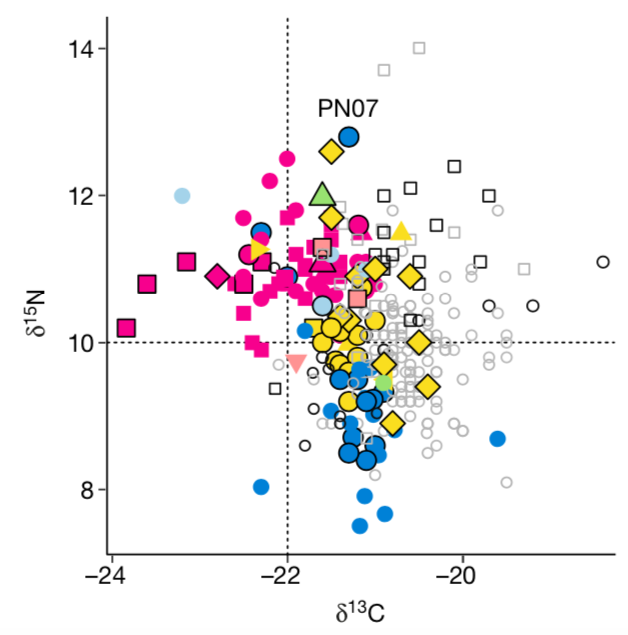
Ganggrabkomplexen bestatteten Individuen zeigten ausserdem
im Vergleich zu den restlichen Bestattungen niedrigere Kohlenstoff-, aber
dafür erhöhte Stickstoffwerte (s. Abb. 4),
die auf einen besseren Zugang zu tierischen Proteinen hinweisen könnten, den
man mit (grosser Vorsicht) einer höheren hierarchischen
Stufe assoziieren könnte. Ausserdem wurde
ersichtlich, dass die ersten Generationen der in Irland bestatteten
neolithischen Bevölkerung ein ähnliches genetisches Signal wie frühe
Ackerbauern aus Spanien zeigen und sich später mit der mesolithischen
Jäger-und-Sammler-Population(en) Irlands vermischt haben.
Abb. 4:
Kohlenstoff- und Stickstoffwerte der irischen und britischen Proben. Pink =
Passage tomb complexes,
gelb = Court tomb, blau = Portal tomb, grün = Linkardstown-Typ,
rosa = natürliche Bestattung, hellblau = nicht klassifizierter Megalith
(Cassidy et al. 2020, 385, Abb. 1b) Einige Proben
für Isotopenanalysen werden je nach Fragestellung aus der mineralischen
Komponente der Knochen, dem Schmelz und Dentin der Zähne entnommen. Hierbei
ist auch die Wahl des Knochens oder des Zahnes essentiell, da diese jeweils
andere Lebensabschnitte und Zeiträume repräsentieren (vgl. Grupe et al. 2015: 68). Dies liegt einerseits an der
Knochenumbaurate, die bei Langknochen langsamer verläuft als bei Rippen oder
anderen flachen Knochen, die eher kürzere Zeiträume vor dem Tod aufzeigen.
Bei den Zähnen ermöglichen die unterschiedlichen Entstehungs- und
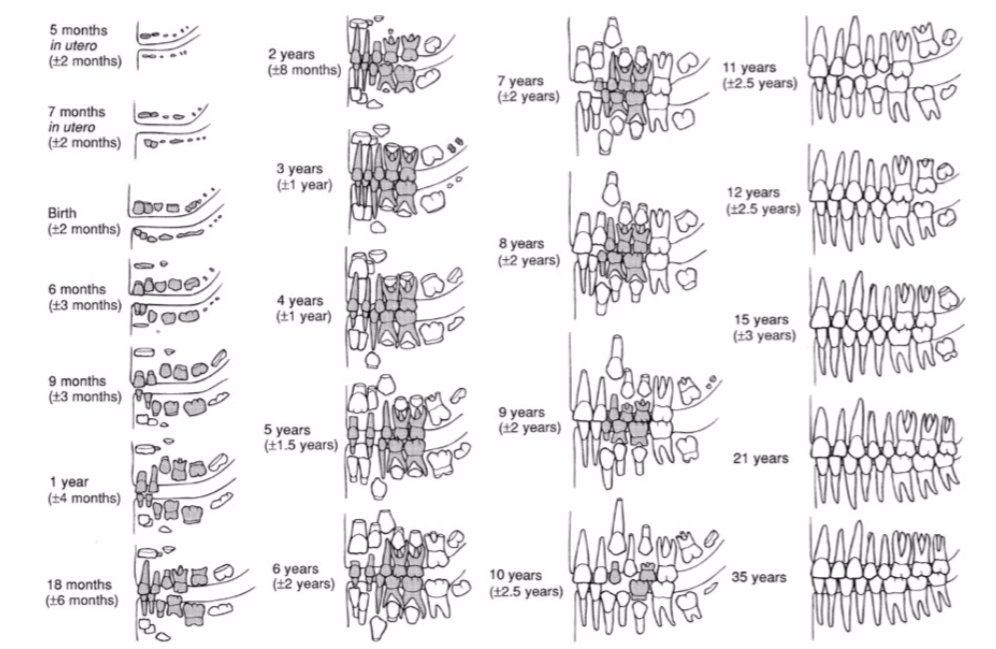
Mineralisationszeitpunkte, verschiedene Zeiträume zu untersuchen (s. Abb. 5), da sich die
mineralische Zusammensetzung, anders als Knochen, nicht mehr verändert und
der Umgebung anpasst (vgl. Grupe et al. 2015: 71);
der Zahnschmelz des ersten Molars gibt beispielsweise Auskunft über die
ersten Lebensjahre, während der dritte Molar die Jugend widerspiegelt (vgl. AlQahtani et al. 2010: 485). Besonders bei Fragen zur
Herkunft und Mobilität im Leben eines Individuums anhand von Sauerstoff- und
Strontiumisotopen ist die Möglichkeit, unterschiedliche Zeiträume zu
untersuchen, sehr wertvoll. Doch auch aus dem Kollagen werden Proben für
Isotopenuntersuchungen entnommen, beispielsweise für
Ernährungsrekonstruktionen anhand von Schwefel-, Kohlenstoff- und Stickstoffisotopen.
Abb. 5:
Entwicklung des Zahnapparats n. D. H. Ubelaker,
Human Skeletal Remains2, 1989. (s. Grupe et al. 2015, 268, Abb. 8.8) Diagenetische
Veränderungen und Reservoir-Effekte erschweren die Interpretation der
Isotopen-Daten, weshalb Referenzdaten aus archäozoologischem
Material und den umgebenden Fundschichten unerlässlich sind (vgl. Burton u.
Price 2013: 309–320). Auch die Erhaltung der Knochen und Zähne spielen eine grosse Rolle, ebenso das Alter und Geschlecht, das bei
der Auswertung miteinbezogen werden sollte, um eine ausführliche Aussage über
die einzelnen Individuen und schliesslich die
untersuchte Population zu ermöglichen. Die hier
erwähnten Isotopenverhältnisse sind stark von den unterschiedlichen
Umwelteinflüssen, der Geologie und dem Klima abhängig und bedürfen einer
genauen und komplexen Auseinandersetzung mit ebendiesen, bevor abschliessende Aussagen gemacht werden können. Dafür braucht es idealerweise Unmengen von
Daten, die sich nicht in den menschlichen Überresten finden, sondern aus der
Umwelt und anderen Überresten, beispielsweise Pollen aus Moorgebieten oder
Nahrungsresten aus zugehörigen Siedlungskontexten, generiert werden müssen. Die
geografische Herkunft sowie die Mobilität der bestatteten Individuen lässt
sich anhand der Isotopenverhältnisse und -werte von Strontium (87Sr/86Sr),
Sauerstoff (∂18O) und Schwefel (∂34S) in Zähnen und Knochen
erörtern, die je nach Geologie und biologisch verfügbaren Signaturen andere
Werte anzeigen. Da Knochen und Dentin sich lebenslang verändern und anpassen,
reflektiert das Isotopenverhältnis im Knochen und Dentin die Werte der
letzten Lebensjahre, während der Schmelz verschiedener Zähne die Signale der
frühen Kindheit und Jugend aufzeigt. Das biologisch
verfügbare Strontium kommt aus der lokalen Geologie und wird von lokalen
Pflanzen und Tieren aufgenommen, bis es schliesslich
zum Endkonsumenten Mensch gelangt und dort anstelle von Kalzium im Bioapatit
der Knochen, im Zahnschmelz und Dentin eingelagert wird (vgl. Grupe et al. 2012: 148–151). Die Strontiumverhältnisse
spiegeln also die geologische Herkunft eines Individuums, bzw. dessen Nahrung
wider (vgl. Grupe et al. 2015: 457, Abb. 10.13;
Knipper 2004). Sauerstoffwerte
dagegen sind stark klimasensitiv und hängen sowohl mit der Temperatur und der
Altitude als auch mit der Entfernung zur Küste und
zum Äquator zusammen (Kontinentaleffekt, Breitengradeffekt). Der Sauerstoff
wird zu mehr als 50% durch das Trinkwasser und die Nahrungsfeuchte aufgenommen
und spiegelt das Ökosystem wider, das damals existierte. Sauerstoff ist in
diesem Sinn ein Marker der ökologisch definierten Herkunft (vgl. Grupe et al. 2012: 145f). Schwefel wird über
die Nahrung aufgenommen und gibt das geologische Signal des Muttergesteins
wieder und ist eng mit dem Niederschlag der Region verknüpft. Es lagert sich
sowohl im Bioapatit als Alternative zum Phosphat als auch in geringeren
Mengen im Kollagen ab, und kann sowohl für die Rekonstruktion der Ernährung
als auch al Herkunftsmarker verwendet werden (vgl. Nehlich
et al. 2014: 15f; Nehlich 2015: 1–17). In
Küstennähe ist auf den sogenannten sea spray effect zu achten, der die pflanzlichen
Schwefelverhältnisse mit marinem Schwefel deutlich erhöht, was jedoch
ermöglicht, marine und terrestrische Ressourcen zu unterscheiden (vgl. Grupe et al. 2012: 147). Um
die im Vorfeld klar definierte Fragestellung zu erörtern und konkrete
Aussagen treffen zu können, benötigt man dringend die lokale Signatur, um
lokale von nicht-lokalen Individuen zu unterscheiden. Referenzen können nicht
mit modernen Daten generiert werden; Dazu sollte(n) die archäologische(n)
Schicht(en), sowie die vorhandene Fauna beprobt werden, um sogenannte
raumzeitliche definierte Isoscapes (isotopic landscapes) zu kartieren,
die erst in den letzten Jahren zunehmend an Qualität gewannen. In Schweden
beispielsweise zeigt sich in den Arbeiten von Sjögren et al. (2009) und
Sjögren (2010), dass die vielen Megalithanlagen in der Region Falbygden im Binnenland einige Individuen enthalten, die
nicht in der Region aufgewachsen sind und erst später nach Falbygden gekommen sind. Auch in anderen Megalithanlagen
in Schweden, wie in der Küstenregion von Bohuslän,
sind nicht-lokale Individuen häufig, zeigen jedoch unterschiedliche Muster
auf. Weitere, klärende Studien zur Differenzierung der lokalen und
nicht-lokalen Individuen, sowie der Variabilität des biologisch verfügbaren
Strontiums in der Region Falbygden und darüber
hinaus werden seit einigen Jahren vermehrt untersucht und zeigen auf, wie
wichtig Referenzdaten zu lokalen Signaturen und rekonstruierte Isoscapes sind (vgl. Blank et al. 2018; 2021). Die genaue
Herkunft von nicht-lokalen Individuen kann nicht abschliessend
definiert werden, da dieselben Signaturen an mehreren Orten vorkommen können.
Ausserdem ist es schwierig, die nicht-lokalen
Signaturen zu interpretieren, denn es gibt nach Sjögren et al. 2009 mehrere
Erklärungen, wie diese zustande kamen, die allesamt andere Interpretationen
zulassen: 1) Einige der lokalen Individuen sind nicht-lokal und kommen aus
einer Region mit ähnlicher Signatur, 2) Einige Individuen sind lokal, aber
aus einer Region, die durch Referenzdaten nicht abgedeckt wurde, 3) Einige
Individuen sind lokal, sind aber etwas mehr in nahen Regionen unterwegs, 4)
Einige Individuen sind lokal, ihre Nahrung stammt aber aus anderen/vielen
verschiedenen Regionen. Eine Kombination mehrerer Isotopen-Signaturen kann
hierbei helfen, mögliche Herkunftsgebiete einzuschränken oder auszuschliessen, wie die Studie von Kador
et al. (2018) beispielsweise am irischen Ganggrabkomplex von Carrowkeel aufzeigt. Die
Ernährung wird anhand der Isotopenwerte von Kohlenstoff (∂13C) und
Stickstoff (∂15N) im Kollagen rekonstruiert, die die Herkunft
pflanzlicher und tierischer Proteine widerspiegeln. Mithilfe der
Kohlenstoffwerte kann zwischen einer Ernährungsweise basierend auf C3- oder
C4-Pflanzen unterschieden werden. Falls auch aquatische Ökosysteme zur
Nahrungsgewinnung beigetragen haben, wird die Interpretation und Aussagekraft
der vorhandenen Ergebnisse jedoch schwieriger. Nicht zu unterschätzen sind
die Trophiestufeneffekte, weshalb im besten Fall
weitere Proben aus der Umgebung auch von archäozoologischen
Funden entnommen werden, um als Referenz ein Nahrungsnetz zu rekonstruieren
(vgl. Grupe et al. 2012: 141–144; 2015: 468, Abb.
10.14a). Anhand der Stickstoffwerte lässt sich eruieren, wie gross der Anteil an tierischen Proteinen (Milchprodukte,
Fleisch) zu Lebzeiten war. Zusammen mit den Kohlenstoffwerten lässt sich sogar
grob zwischen Ackerbau und Viehzucht, und weiter zwischen Milch- und
Fleischwirtschaft unterscheiden. Die Ergebnisse liefern wertvolle
Informationen zum Status eines Individuums oder aber auch der Verteilung,
respektive den Zugang zu tierischen Proteinen und der Ernährungsweise(n)
innerhalb der untersuchten Population (s. Abb. 4), und kann uns Hinweise auf
die Wirtschaftsweise und Hierarchie der untersuchten Gesellschaft geben.
Schwefelisotopenverhältnisse können zur Unterscheidung von marinen und terrestrischen
Ressourcen hinzugezogen werden (s. oben). Allerdings können die
ernährungsspezifischen Isotopenwerte stark von gesundheitlichen Faktoren
sowie bei Frauen und Kindern vom Stillen abhängig sein. 2.3.3 Die Isotopen-Untersuchungen in der Schweiz Die Bestattungen
des 2012/13 gegrabenen spätneolithischen Dolmen von Oberbipp im Kanton Bern
konnten dank ihrer guten Erhaltung bioarchäologisch untersucht werden; nebst
einer detaillierten 14C-Datierung anhand der Oberschenkelknochen (ca.
3350–2950 u. 2900–2650 v. Chr.) wurden auch genetische Untersuchungen und
mehrere Isotopenanalysen durchgeführt. Von den mind. 42 bestatteten
Individuen, die nicht mehr oder nur teilweise im anatomisch korrekten Verband
gefunden wurden, konnten etwa gleich viele männliche als auch weibliche
Individuen sowohl morphologisch als auch, wenn möglich, genetisch bestimmt
werden. Bei einigen Individuen war eine Geschlechtsbestimmung nicht eindeutig
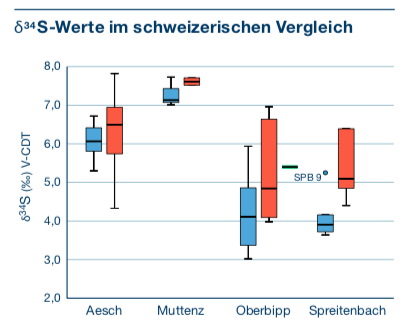
möglich. Die Schwefelwerte (s. Abb. 6) deuten darauf hin,
dass einige Individuen beider Geschlechter nicht-lokal aufgewachsen sind oder
sich von nicht-lokaler Nahrung ernährt haben. Referenzdaten lieferten mehrere
Tierknochen aus der Umgebung Oberbipp und aus den etwa zeitgleich datierten
Fundschichten der Seeufersiedlung im entfernt gelegenen Twann, Bern. Da die
Signaturen ungefähr dem erwarteten Wert für das Schweizer Mitteland
entsprechen, können sie lediglich als ortsfremde Individuen angesprochen
werden. Die weiblichen Individuen haben interessanterweise eine grössere Streuung der Schwefelwerte, was darauf hinweisen
kann, dass tendenziell Frauen aus unterschiedlichen Regionen nach Oberbipp
kamen und die Männer aus einer homogenen Region und ortstreu
waren. Es könnte sich daher um eine patrilokale
Gesellschaftsstruktur handeln, was allerdings nur durch weitere Daten, wie
beispielsweise Strontium- und Sauerstoffisotopie,
genauer untersucht werden kann.
Abb.
6: Boxplot der Schwefelisotopenwerte verschiedener neolithischer Schweizer
Fundorte: Aesch BL, Muttenz BL, Oberbipp BE, Spreitenbach AG. Rot = weibliche
Individuen, blau = männliche Individuen, grün = unbestimmt (Lösch et al.
2020, 219, Abb. 14) Die
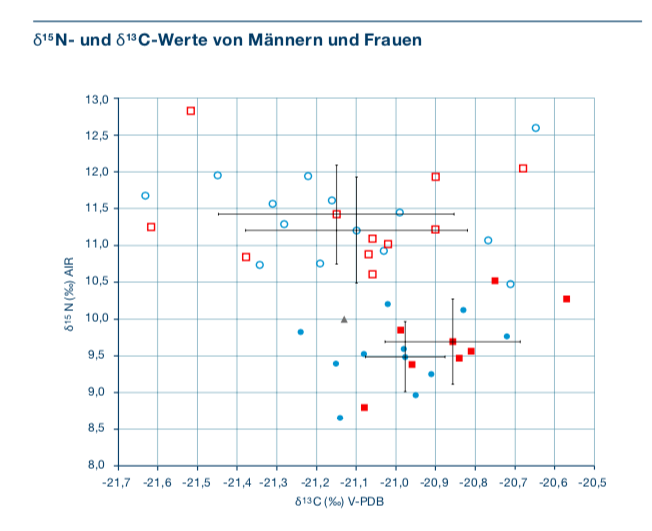
Kohlenstoff- und Stickstoffwerte (s. Abb. 7) zeigen, dass
sich die Bestatteten ortstypisch ernährten und kein signifikanter Unterschied
zwischen den Geschlechtern in Bezug auf den Zugang zu tierischen Proteinen
bestand. Die genetischen Analysen ergaben, dass es sich bei einigen
Individuen um Brüder, Väter und Söhne handelt, die über drei Generationen
miteinander verwandt sind, während bei den weiblichen Individuen keine
verwandtschaftlichen Bande nachgewiesen werden konnten, was wiederum eher für
eine patrilokale Organisation spricht.
Abb.
7: Kohlenstoff- und Stickstoffwerte der Individuen von Oberbipp BE
(ausgefüllt) im Vergleich zu Aesch BL (leer) inklusive Mittelwerte mit
Standardabweichung. Rot = weibliche Individuen, blau = männliche Individuen,
grau = unbestimmt (Lösch et al. 2020, 218, Abb. 12) Insgesamt vermuten Lösch et al., dass es sich bei
den im Dolmen von Oberbipp bestatteten Individuen nur um einen ausgewählten
Teil der Bevölkerung handelt, vermutlich einer höheren Elite. Dabei sind
vermutlich tendenziell mehr Männer als Frauen bestattet worden, die jedoch
keinen signifikant unterschiedlichen Zugang zu proteinreicher Nahrung hatten
und deshalb keine soziale Stratifizierung in Bezug auf die
geschlechterspezifische Ernährung vermutet wird. Aufgrund der
Schwefelisotopenwerte und der genetischen Verwandtschaft zwischen männlichen
Individuen, handelte es sich eventuell um eine patrilokale
Gesellschaft, die hier bestattet wurde. Wie im Eingangszitat von Alain Gallay
dargestellt, lässt sich die Spezies Mensch, weder als Individuum noch als
Teil einer Gemeinschaft, nicht anhand einer einzigen der vielen Teilaspekte
beschreiben. So stellen die bioarchäologischen Ergebnisse nur einen Bruchteil
der Informationen dar, die für eine Annäherung an die Realität nötig sind.
Weitere Erkenntnisse aus dem ethnoarchäologischen Bereich, dem materiellen
Kontext sowie der Sozialanthropologie und der Kulturwissenschaften müssen
hinzugezogen werden. Dieser interdisziplinäre Ansatz ermöglicht es, den Versuch
einer Rekonstruktion der megalithischen Gesellschaften im europäischen
Neolithikum zu wagen. Die Kollektivbestattungen, die aus historischen und zeitgenössischen
Beispielen aus Indonesien von den Toraja bekannt
sind, stellen dabei eine Möglichkeit dar, wie megalithischen Gesellschaften
sozial organisiert sein können und wie sich eben dies im funeralen Kontext
zeigt. Allerdings sind die Vergleiche mit Vorsicht zu behandeln, da
beispielsweise bei den Toraja die traditionellen
Praktiken und Riten zwar erhalten, aber auch modernisiert und verändert
wurden, und es in Bezug auf die räumliche Organisation zwischen den ‹Häusern
der Lebenden› und den ‹Häusern der Toten› drastische Unterschiede zwischen
den megalithischen Gesellschaften Indonesiens gibt (vgl. Jeunesse u. Denaire 2018: 86, 93). Dennoch wird an diesem Vergleich
ersichtlich, dass die biologische Verwandtschaft (biological
kinship) eine essentielle Rolle bei den
Kollektivbestattungen sowie in der sozialen Organisation spielt; die
Abstammung, die ancestry, hat grosses
Gewicht, wobei ein Individuum jedoch aussuchen kann, ob es bei der eigenen
Bestattung im maternalen, paternalen oder maritalen Kollektivgrab beigesetzt werden möchte (vgl.
Jeunesse u. Denaire 2018: 86f, 96). Zwischen den
verschiedenen Gruppen besteht eine Konkurrenz um Prestige, die durch
ostentative Praktiken dargestellt wird, beispielsweise durch
Bestattungszeremonien und dem megalithischen Zyklus. Diese Gruppen werden
durch lineages definiert; hierbei handelt es sich
jeweils um eine unilineare Gruppe, deren Abstammung entweder der männlichen
oder der weiblichen Linie folgt, aber nicht beiden. Die Abstammung spielt
eine wichtige Rolle in der sozialen Organisation der Gesellschaft. Davon zu
unterscheiden ist der Begriff der lineage society, die eine Gesellschaft beschreibt, die ausschliesslich über Abstammungslinien definiert wird
(vgl. Testart 2014: 333). Jeunesse und Denaire argumentieren, dass
die neolithischen Megalithen und die damit assoziierten Gesellschaften auf
ähnliche Art und Weise organisiert gewesen sein könnten (vgl. Jeunesse u. Denaire 2018: 87).
Dies stellt jedoch nur einen kurzen Exkurs in die sehr komplex
organisierten, Megalithen-erbauenden Gesellschaften wie der Toraja und Niha, der Merina und der Rapanui dar, die
als ethnoarchäologische Vergleiche herangezogen werden können. Die bioarchäologischen Erkenntnisse stützen die verschiedenen
Beispiele sozialer Strukturen, die aus dem ethnoarchäologischen Vergleich
ersichtlich werden. So sehen wir zwar die biologische Verwandtschaft in den
Analyseergebnissen, jedoch wird erst aus dem archäologischen Kontext auch
eine rein soziale Verbindung zwischen Individuen oder Gruppen deutlich; dass
die biologische Reproduktion eine grosse Rolle in
den neolithischen Gesellschaften spielt, ist aus unzähligen
Bestattungskontexten ersichtlich, sowohl bei megalithischen als auch bei
nicht-megalithischen Kollektivbestattungen, wie beispielsweise der Totenhütte
von Benzingerode (vgl. Meyer et al. 2008; 2012a:
14ff; 2012b: 147–150). Doch auch die soziale Reproduktion, die teilweise von der
biologischen abhängig ist, darf nicht vergessen werden (vgl. Meyer et al.
2012a: 11–14, 21). Ein wichtiger Aspekt der sozialen Reproduktion ist
besonders die Erinnerung; sowohl auf der individuellen Ebene wie auch auf der
Ebene der Gemeinschaft. Dabei besteht innerhalb jeder Gemeinschaft ein
«kollektives Gedächtnis», das aus dem «kommunikativen» (mündliche
Überlieferungen, oral history) und dem «kulturellen
Gedächtnis» (archäologischer u. schriftlicher Nachlass) besteht (vgl. Assmann
2013: 35–37, 45, 50–53). 3.1
Das kulturelle Gedächtnis nach Assmann Unter dem Begriff «kulturelles Gedächtnis» versteht Jan Assmann
(*1938) einen Teil der Aussendimension des
menschlichen Gedächtnisses. Dabei handelt es sich beim menschlichen
Gedächtnis nicht nur um eine innere Struktur, die von einem Menschen selbst
bestimmt wird, sondern um ein Konzept, das auch von den Mitmenschen und der
Gesellschaft, in der sich der Mensch befindet, strukturiert wird. So sind
Inhalt, Organisation und Länge des Erinnerns von gesellschaftlichen und
kulturellen Faktoren abhängig (vgl. Assmann 2013: 19–21). Nach Maurice
Halbwachs (1877–1945) kann diese Aussendimension
des menschlichen Gedächtnisses in vier Bereiche unterteilt werden: Das
mimetische Gedächtnis (Handeln, Nachmachen), das Gedächtnis der Dinge
(Materielles, das ein Bild der Entität widerspiegelt und Erinnerungen
beinhaltet), das kommunikative Gedächtnis (Sprache, Kommunikation, zeitlich
an Generationen gebunden) und das kulturelle Gedächtnis (Überlieferung des
Sinns, Mythen), wobei die ersten drei Bereiche auch in den «Raum» des
kulturellen Gedächtnisses einfliessen. Im
kulturellen Gedächtnis sind nicht nur der Zweck, sondern auch der Sinn von grosser Bedeutung. Dabei geht es immer auch in gewisser
Weise um die Identität, die bei Individuen und Gemeinschaften eine
Unterteilung in «Wir», «Ihr» und «Ich» zulässt. Das «Wir» basiert stark auf
einer kontinuierlichen, sich wiederholenden Reproduktion der gemeinsamen
Werte der untersuchten Gesellschaft. Die Erinnerung an den gemeinsamen
Vergangenheitsbezug und die in jeder Gesellschaft sichtbare Traditionsbildung
(kulturelle Kontinuierung), bildet eine konnektive Struktur, die sowohl auf einer sozialen wie
auch auf einer raum-zeitlichen Ebene verbindet und eine Orientierung
gewährleistet. In schriftlosen Gesellschaften zeigt sich dies in Form von
mündlichen Überlieferungen (und deren spezialisierten Träger*innen, vgl.
Assmann 2013: 54f), sowie bildlichen und symbolischen Darstellungen, sowohl
im materiellen als auch im immateriellen Kontext anhand von Riten, Festen,
Totengedenken und Monumenten (vgl. Assmann 2013: 16f, 56–63). In der Archäologie treffen wir diese Wiederholung in Form von
erkennbaren Mustern und Elementen an, die dann als zusammengehörendes
Kollektiv angesprochen werden können; ich verweise auf den kontroversen
Begriff der ‹Kultur›, der lange Zeit gebräuchlich war. Es gibt zwar
verbindende Elemente und Monumente zwischen den einzelnen, mit megalithischen
Strukturen assoziierten Individuen und Gemeinschaften, doch kann nicht von
einer einzelnen Gesellschaft ausgegangen werden. Es handelt sich im
europäischen Neolithikum vielmehr um verschiedene Gesellschaften, die sich in
Anbetracht des Konzepts des kulturellen Gedächtnisses ähneln können, indem
sie zur Erinnerung und Identitätsvergegenwärtigung megalithische Monumente
errichteten. Innerhalb der einzelnen Gesellschaften wird, gestützt durch
biologische Erkenntnisse, eine entfernte gemeinsame Vergangenheit geteilt,
die so zum Ausdruck gebracht wird. Allerdings soll darauf hingewiesen werden,
dass Unterschiede bestehen können, die sich uns im materiellen Kontext nicht
zeigen, sondern nur anhand von ethnoarchäologischen Vergleichen ersichtlich
sind. Das Konzept des kulturellen Gedächtnisses ermöglicht es uns jedoch,
eine abstrahierte Perspektive auf die neolithischen Gesellschaften zu
erkunden, die wiederum neue Ansätze der zukünftigen Forschung aufzeigen kann. Die Untersuchung von Kollektivbestattungen in megalithischen Anlagen,
wie Dolmen, Gang- und Passgräber, die im Neolithikum in Nord- und Westeuropa
für Bestattungen über längere Zeiträume verwendet wurden, profitieren von den
bioarchäologischen Analysen. Zusammen mit einer guten Dokumentation und
umfassenden anthropologischen Untersuchung lassen sich anhand von aDNA-Analysen bereits vermutete, im Befund ersichtliche
Beziehungen und Verwandtschaften weiter untersuchen, während Isotopenanalysen
Einblicke in die Herkunft, Mobilität, Verbindung und Ernährung der Individuen
ermöglichen. Allerdings zeigen die genetischen Marker nur die biologische,
nicht aber die soziale Verwandtschaft an. So lassen sich nur im Zusammenhang
mit dem archäologischen Kontext Aussagen formulieren, die über die
biologischen Beziehungen (individuelle Ebene) hinausgehen und soziale
Verbindungen (Gemeinschaftsebene) sichtbar machen können. Betrachtet man nun diese verschiedenen bioarchäologischen Studien
megalithischer und nicht-megalithischer Kollektivbestattungen, wird
ersichtlich, dass es sich um verschiedene Gruppen oder Populationen handelt,
die gesellschaftlich ähnlich, aber nicht einheitlich organisiert gewesen sein
könnten. Die biologische Verwandtschaft konnte allerdings in allen genetisch
untersuchten Populationen nachgewiesen werden, so dass die biologische
Reproduktion nebst der sozialen Reproduktion, die aus den archäologischen und
ethnoarchäologischen Betrachtungen ersichtlich ist, eine wichtige, wenn nicht
gar essentielle und verknüpfende, Komponente der diversen megalithischen und
zeitgleichen nicht-megalithischen Gesellschaften darstellt. Auch eine
mesolithische Komponente konnte bei den populationsgenetisch untersuchten
Individuen festgestellt werden, was die von Alain Testart vorgeschlagene
These unterstützt, dass das Errichten megalithischer Bauten einen Ursprung in
den mesolithischen Jäger-und-Sammler-Gesellschaften haben und durch den
Prozess der «Neolithisierung» aufgekommen sein könnte (vgl. Cauwe 2016: 219). Die soziale Verbundenheit, die weder genetisch noch biochemisch
nachgewiesen werden kann, wird jedoch in den erwähnten Grabkontexten zwischen
einzelnen Individuen deutlich. Diese selbstbestimmten Netzwerke waren für die
neolithischen Gesellschaften, wie auch bei einigen zeitgenössischen
megalithischen Gesellschaften, unerlässlich. Dazu haben viele dieser
miteinander verglichenen Gesellschaften gemein, dass megalithische Monumente,
deren längerer Nutzungszeitraum und die Dualität von Leben und Tod, sowie
deren Verbindung, wichtige Aspekte innerhalb der Gesellschaft verkörpern.
Dies ist unter anderem mit Jan Assmann’s Konzept
des kulturellen Gedächtnisses vereinbar, was uns eine erweiterte Perspektive
in immaterielle Aspekte erlaubt, die mit dem materiellen Kontext verbunden
werden kann. Acsádi u. Neméskeri 1970 AlQahtani et al. 2010 Alt et al. 2016 Assmann 2013 Blank et al. 2018 Blank et al. 2021 Boulestin 2016 Burton u. Price
2013 Cassidy et al.
2020 Cauwe 2014 Cauwe 2016 Ferembach et al. 1979 Gallay 2011 Grupe et al. 2012 Grupe et al. 2015 Jeunesse u. Denaire 2018 Kador et al. 2018 Knipper 2004 Le Roy 2018 Lösch et al. 2020 Meyer et al. 2008 Meyer et al. 2012a Meyer et al.
2012b Nehlich et al. 2014 Nehlich 2015 Perréard Lopreno 2014 Sánchez-Quinto et al. 2019 Schwegler 2016 Sjögren et al. 2009 Sjögren 2010 Steimer-Herbet 2018 Testart 2014 Weiss-Krejci 2018 |
|
|
|
|
|
|